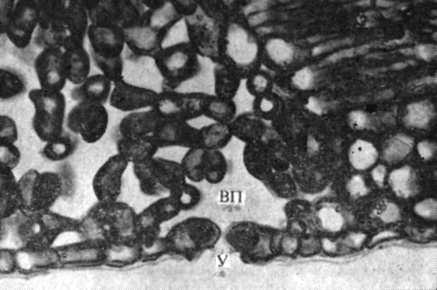
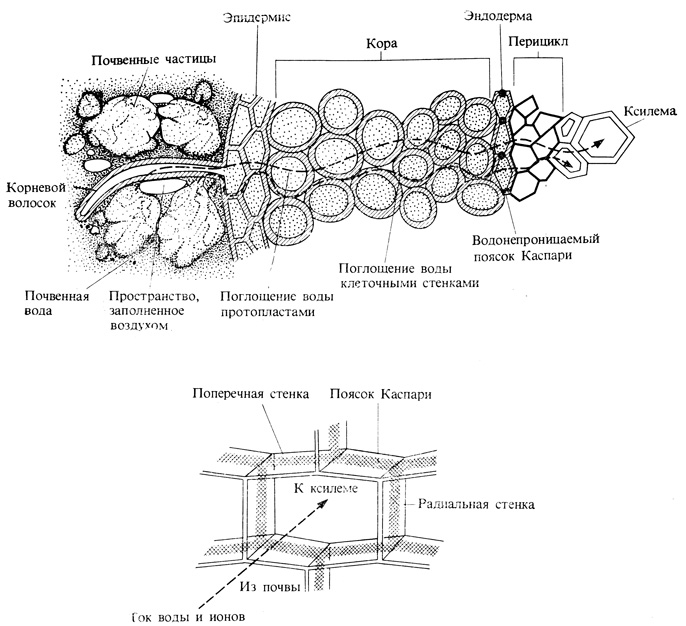
Вода движется в растении по градиенту водного потенциала. Вода, поглощенная корневыми волосками и другими клетками эпидермиса, из клеток внешней части корня перемещается к ксилеме, занимающей центральную часть корня (рис. 6.8). Главным путем диффузии воды во внешней части корня служит апопласт - непрерывная совокупность клеточных стенок. Однако в эндодерме (цилиндрическом слое клеток, окружающем проводящую ткань) свободная диффузия по клеточным стенкам наталкивается на преграду - водонепроницаемый пробковый слой пояска Каспари. Вода должна изменить здесь свой путь и пройти сквозь мембрану и протопласт клеток эндодермы, играющей, таким образом, роль осмотического барьера между корой корня и его центральным цилиндром. У однодольных пробковеют также и внутренние тангенциальные стенки клеток, но эти стенки пронизаны порами, по которым, как по каналам, может проходить вода.
По ксилеме вода поднимается в надземные части растения. Ксилема состоит из нескольких типов клеток. Вода движется в ней главным образом по сосудам и трахеидам (рис. 2.6 и 6.9). И те и другие клетки прекрасно приспособлены для этой цели: они вытянуты в длину, лишены живого содержимого и внутри полые, т. е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огромную разность давлений, возникающую при подъеме воды к вершинам высоких деревьев. Торцевые, а иногда и боковые стенки члеников сосудов перфорированы; сосуды, состоящие из соединенных конец в конец члеников, образуют длинные трубки, по которым легко проходит вода с растворенными в ней минеральными веществами. В трахеидах нет перфораций, и вода, для того чтобы попасть из одной трахеиды в другую, должна пройти через их торцевые стенки; однако трахеиды - очень длинные клетки, а потому и эта конструкция достаточно хорошо приспособлена для проведения воды. У цветковых растений есть и сосуды, и трахеиды; у более при-митивных форм сосудов, как правило, нет.
Листовые жилки, состоящие из тяжей ксилемы и флоэмы, образуют в листе настолько густую сеть, что любая его клетка оказывается достаточно близко от источника воды (рис. 3.9). Из ксилемы вода диффундирует в стенки клеток мезофилла. Таким образом, вода в жидкой фазе заполняет весь путь от почвы - через корень и стебель - до клеток мезофилла в листе. Суммарный поток воды направлен всегда в сторону меньшего водного потенциала, т. е. ψ максимален в почве, несколько ниже в клетках корня и самый низкий в клетках, примыкающих к эпидермимису листа. Малая величина ψ в этих последних клетках объясняется главным образом испарением воды с поверхности листа, т. е. транспирацией (которой посвящен следующий раздел). Табл. 6.2 дает представление о градиентах, обусловливающих движение воды в растении от ее поступления из почвы до испарения в атмосферу.
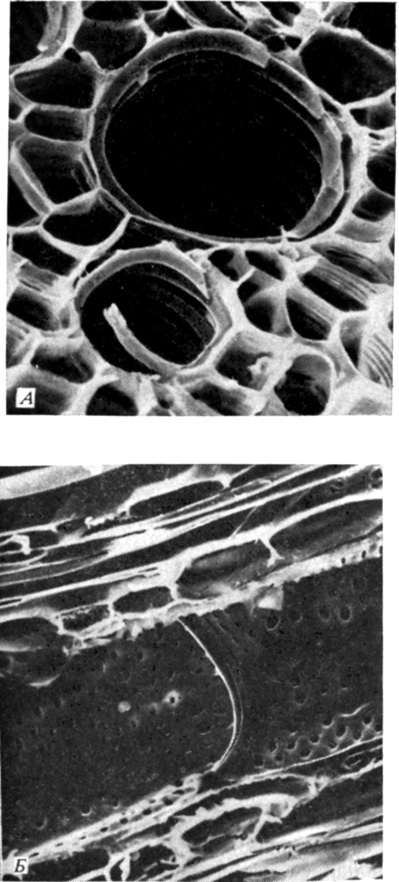

Таблица 6.2 Примерные значения водного потенциала (ψ) и разности водных потенциалов (Δψ) для гипотетической системы почва - растение - воздух. [Предполагаемые условия - сравнительно небольшое дерево, хорошо увлажненная почва, относительная влажность воздуха около 50% при 22 °С (ψ=-1000 бар) 1)]
( 1) R. G. S. Bidwell. 1974. Plant Physiology, New York, Macmillan. )
Вода переходит из растения в окружающий воздух главным образом в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства, и каждая клетка мезофилла хотя бы одной своей стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, и часть этих паров теряется - выходит наружу. Поскольку у большинства растений клетки эпидермиса покрыты воскообразной водонепроницаемой кутикулой, водяные пары выходят из листа в атмосферу главным образом через устьица (рис. 6.10).